|
Лидский П. Механизмы нуклеоцитоплазматического транспорта
Курсовая работа студента 4-го курса кафедры вирусологии Биологического
факультета МГУ. Москва, 2001
Авторские права сохранены. Любое копирование данного текста и/или его
фрагментов без разрешения автора запрещено и преследуется в соответствии
с действующим законодательством РФ.
Содержание В начало...
Экспериментальные подходы
Для осмысления каких-либо научных данных, часто необходимо понимать, каким образом эти данные были получены. Поскольку исследователи ядерно-цитоплазматического транспорта часто используют довольно необычные объекты и эксперименты, имеет смысл описать некоторые из них.
 |
| Рис 11. Ооциты
|
Во-первых, эксперименты проводятся на ооцитах Xenopus laevis. В эту огромную клетку (Рис. 11) микроинъецируют субстрат, после чего ядро отделяют от цитоплазмы, а накопление субстрата в той или иной фракции определяют по включению радиоактивной метки или, в последние годы, проводят иммунноблотинг. На ооцитах, например, была сделана пионерская работа, доказавшая независимость различных видов транспорта РНК [Jarmolowski, A. et. al. 1994].
Второй тип экспериментов ставится на клеточных культурах, обработанных дигитонином. Это вещество повреждает внешнюю мембрану клетки, но оставляет интактной ядерную оболочку [Adam et. al. 1990]. Тем самым можно вымыть из такой клетки все цитоплазматические компоненты транспортных систем и исследовать транспорт различных субстратов, добавляя препараты белков, подозреваемых на функцию транспортинов для данных субстратов. Если, к примеру, флуоресцентный субстрат накапливается в ядре (Рис 12, верхняя половина), значит подозреваемый белок отвечает за его импорт. С другой стороны, можно истощать цитоплазматические экстракты тех же ооцитов Xenopus на разных адсорбентах и тестировать их после этого на возможность индуцировать транспорт тех же субстратов. Например, если истощение экстракта на никель - агарозе ингибирует импорт NLS-белков, значит, с данным адсорбентом связывается фактор, принимающий участие в этом процессе (так был найден импортин-alfa) [Gorlich et. al.1994].
 |
| Рис 12. |
Еще один тип экспериментов производится на дрожжах с помощью молекулярно-генетических методов (Рис 13. Флуоресцентный анализ, красное- миозин-подобный белок, синее- ДНК). На этом объекте было получено большое количество сведений относительно строения ядерных пор и действия транспортинов. К сожалению, транспортные системы дрожжей довольно значительно отличаются от таковых многоклеточных.
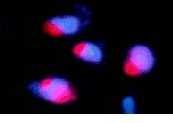 |
| Рис 13. Ядра сферопластов. |
Наконец, для исследования физико-химических свойств ядерных пор, используется целый спектр биофизических методов. Среди них можно упомянуть метод силовой атомной микроскопии (AFM; atomic force microscopy), позволяющий наблюдать состояние ядерной поры в растворе. Это позволяет исследовать действие различных факторов на диаметр ядерной поры и ее высоту (Рис 14) [по T. Danker, H. Oberleithner, 2000].
 |
| Рис 14. |
Еще один необычных биофизический метод - так называемый метод ядерных песочных (nuclear hourglass) часов. Этот метод позволяет определять электропроводимость ядерных пор более точно для больших ядер, чем метод с использованием микроэлектродов. Суть метода заключается в закреплении ядра в центре сосуда, имеющего форму песочных часов, и измерении сопротивления между верхним и нижним отделениями (Рис 15) [по T. Danker, H. Oberleithner, 2000].
 |
| Рис 15. |
Заключение
Нуклеоцитоплазматический транспорт требует весьма значительных энергетических затрат. В этом процессе принимают участие многие десятки белков, иногда очень многочисленных в эукариотической клетке. Какие же выгоды извлекает клетка из компартментализации, и какие причины обусловили ее появление в эволюции?
Сначала попытаемся дать ответ на второй вопрос. Во-первых, жесткое разделение процессов транскрипции и трансляции, которое появилось у эукариот, позволило принципиально усложнить и усовершенствовать системы процессинга РНК. Действительно, то широкое распространение, которое получила интрон-экзонная схема строения генов в эукариотических клетках, невозможна в случае прокариот, где системам процессинга пришлось бы конкурировать за РНК с рибосомами. Еще раз хочется подчеркнуть, что системы ядерного экспорта РНК селективны по отношению к процессированным и непроцессированным субстратам, что еще раз подтверждает необходимость компартментализации для процессинга.
Во-вторых, возникновение органеллы приспособленной для хранения генетического материала привело к повышению стабильности этого материала. А это позволило значительно увеличить размеры геномов. Действительно, геномы прокариот и эукариот по объему отличаются на порядки.
Какие еще преимущества дает компартментализация? Можно высказать еще, по меньшей мере, два соображения. Первое состоит в том, что возникновение барьера между цитоплазмой и областью хранения генетического материала, позволило клетке выработать качественно новые регуляторные механизмы. Наряду с увеличением размеров генома, это вероятно, послужило мощным стимулом для развития многоклеточности.
Второе, более частное, соображение состоит в том, что защита генетического материала дополнительной оболочкой может усложнить жизнь ДНК-содержащим вирусам, как это имеет место, например, в случае некоторых представителей Lentiviridae, размножающихся только в делящихся клетках. Можно даже предположить, что на заре эволюции, этот фактор давал значительные селективные преимущества. Но вирусы не остались в долгу. Эти организмы не только изобрели механизмы, позволяющие им попадать в ядро и выходить из него, используя хозяйские транспортные системы, но и эксплуатировать эти системы для регуляции собственного цикла. Появление обратной транскрипции у вирусов (Retroviridae, Hepadnaviridae) также могло произойти как приспособление к существованию в эукариотах. Действительно, поскольку генетический материал вируса экспортируется из ядра в качестве РНК, оказывается возможным использовать клеточный механизм транспорта. Таким образом, для репликации ретровирусов не нужно ни разрушение ядра, ни кодирование специальной вирусной транспортной системы.
Далее...
Написать комментарий
|
