|
Мончева П.А., Данова С.Т., Гочева Я., Иванова И.В. Биологический факультет Софийского университета им. Св. Кл. Охридского, Болгария
В начало...
(Окончание)
Как было показано выше, исходный штамм S.hygroscopicus 155 продуцирует антибиотический комплекс, основными компонентами которого являются антибиотики пандавир (нигерицин) и азаломицин В. Ранее нами было установлено, что неактивный вариант родительского штамма S.hygroscopicus 155-0 высокочувствителен к пандавиру, внесенному извне в жидкую питательную среду [18]. В литературе высказаны предположения о регуляторной функции антибиотиков по отношению к их продуцентам. Было исследовано влияние пандавира и азаломицина (нанесенных на бумажные диски) на формирование воздушного мицелия у штамма S.hygroscopicus 155-0. Пандавир, добавленный в среду, не содержащую Са2+, индуцировал формирование воздушного мицелия, в то же время наблюдалось подавление развития штамма в непосредственной близости к диску. Азаломицин также индуцировал образование воздушного мицелия, но без подавления развития штамма. Интересен тот факт, что антибиотик монензин, близкий по структуре к нигерицину (пандавиру), также индуцировал образование воздушного мицелия, в то время как салиномицин не проявлял такого действия. Монензин и салиномицин, а также пандавир являются полиэфирными антибиотиками с ионофорными свойствами. Было исследовано одновременное действие Са2+, пандавира и азаломицина и модуляторов Са2+-сигналов (в этом случае каждый диск был пропитан тремя соединениями одновременно). Наблюдалось формирование обильного воздушного мицелия и в то же время снимался подавляющий эффект нифедипина в присутствии двух антибиотиков, а также эффект верапамила и дилтиазема при комбинации с азаломицином В. Ранее нами было установлено аналогичное стимулирующее действие экзогенно добавленного собственного антибиотика на дифференциацию штамма Streptomyces albogriseolus 444 (также продуцент нигерицина) [9]. Это действие усиливалось в присутствии Са2+ в среде. Таким образом, можно предположить, что собственные антибиотики выполняют регуляторную функцию в микроорганизмах-продуцентах и что эта их функция экспрессируется через Са2+-сигнальную систему.
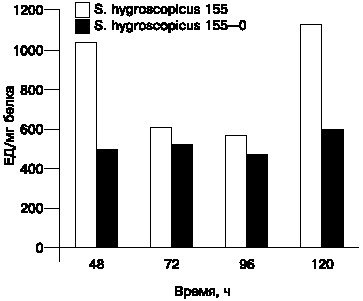
В связи с высказанным рядом авторов предположением о роли внеклеточных протеаз в процессе дифференциации стрептомицетов и на основе проведенных нами исследований с S.albogriseolus 444 [14], была изучена активность внеклеточных протеаз на разных этапах культивирования S.hygroscopicus 155 и его неактивного варианта. Посредством использования различных протеазных ингибиторов было установлено, что S.hygroscopicus 155 (подобно S.albogriseolus 444) продуцирует протеолитический комплекс, включающий металлопротеазы и трипсинподобные протеазы (результаты не приводятся). Была определена общая протеазная (казеиназная) активность S.hygroscopicus 155 и S.hygroscopicus 155-0. Результаты представлены на рис. 1. Видно, что общая протеолитическая активность неактивного штамма находится на более низком уровне по сравнению с родительским штаммом. Следует отметить, что среда, в которой культивировались штаммы, не содержала субстрата протеаз, т.е. по всей вероятности эти протеазы являются конститутивными. Laluce и Molinari [19] также выявили внеклеточную казеиназную активность Streptomyces aureofaciens при наличии и отсутствии субстрата.
 |
 |
| Рис. 1. Протеолитическая (казеинолитическая) активность S.hygroscopicus 155 и S.hygroscopicus 155-0. |
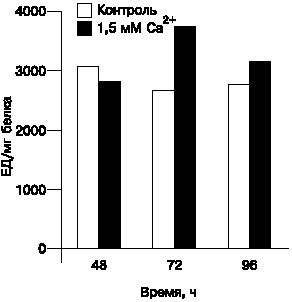
Рис. 2. Влияние Са2+ на протеолитическую (казеинолитическую) активность S.hygroscopicus 155. |
 |
 |
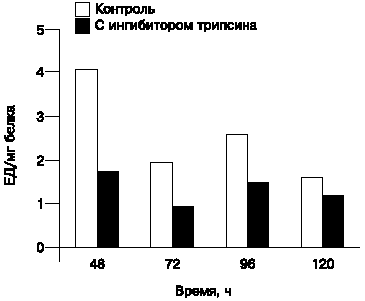
| Рис. 3. Протеолитическая активность (субстрат БАПНА) штамма S.hygroscopicus 155 в присутствии ингибитора трипсинподобных протеаз. |
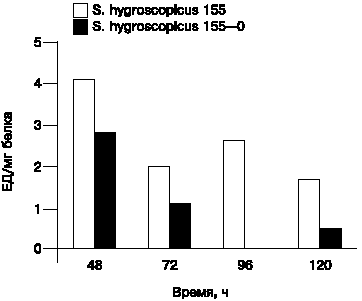
Рис. 4. Протеолитическая активность (субстрат БАПНА) штаммов S.hygroscopicus 155 и S.hygroscopicus 155-0. |
Далее было показано, что Са2+ в концентрации 1,5 мМ стимулирует протеазную активность штаммов через 48 ч после начала культивирования (рис. 2). Возможно, что этот эффект связан с влиянием кальция на дифференциацию культур. Протеолитическая активность регулируется не только посредством синтеза новых молекул, но также и посредством их активирования [20]. В последние годы получен ряд доказательств о поддержании высокого уровня общей внутриклеточной протеазной активности в присутствии кальциевых ионов во время споруляции бактерий рода Bacillus [21, 22]. Эффект кальция объяснялся и его стабилизирующим действием на активность сериновых протеаз [23].
Была исследована субстратная специфичность секретированных протеаз только относительно БАПНА (специфический субстрат трипсинподобных протеаз) в условиях добавления трипсинового ингибитора in vitro. Было установлено, что добавление трипсинового ингибитора к реакционной смеси ингибирует активность протеаз, синтезированных штаммом S.hygroscopicus 155 (рис. 3).
Проведено сравнение активности трипсинподобных протеаз родительского штамма и его неактивного варианта (рис. 4). Активность была более высокой у родительского штамма. Это дает основание искать связь между синтезом внеклеточных протеаз, дифференциацией и антибиотикообразованием. В литературе существуют данные, показывающие прямо пропорциональную зависимость между антибиотикообразованием и внеклеточной протеолитической активностью [24].
Исходя из полученных результатов можно предположить, что ионы кальция индуцируют (стимулируют) процессы дифференциации у S.hygroscopicus 155-0 и S.hygroscopicus 155. Блокирование их индуцирующего действия модуляторами Са2+-каналов подтверждает их роль в процессах дифференциации. Возможно функционирование кальмодулинподобных протеинов с кальций-связывающими функциями.
Индуцирующая роль продуцируемых родительским штаммом антибиотиков на дифференциацию S.hygroscopicus 155-0, а также усиление их действия в присутствии Са2+ дают основание предположить, что они являются регуляторами дифференциации, и эта их функция экспрессируется через Са2+-сигнальную систему.
Низкая протеолитическая активность неактивного, недифференцированного варианта родительского штамма позволяет искать связь между дифференциацией, антибиотикообразованием и внеклеточной протеолитической активностью.
Данная работа выполнена на основании проекта К-610/96, финансированного Фондом научных исследований МОНТ Болгарии.
АНТИБИОТИКИ И ХИМИОТЕРАПИЯ, 2000-N2, стр. 10-14
1. Khokhlov A.S., Anisova L.N., Tovarova I.I. et al. Z Allg Mikrobiol 1973; 13: 647655.
2. Szabo G., Valyi-Nagy T., Vitalis S. Acta Biol Hung 1967; 18: 237-243.
3. Shapiro S. In: Regulation of Secondary Metabolism in Actinomycetes/Ed by S. Shapiro. Boca Raton 1989; 149-153.
4. Rowley D.B., Levinson H.S. J Bacteriol 1967; 93: 1017-1022.
5. Sacks L.E. In: Spores V/Ed by H.O. Halvoson, R. Hanson, L.L. Campbell. Washington 1972; 437-443.
6. Kamat A.S., Lewis N.F., Pradham D.S. Microbios 1985; 44: 33-44.
7. Natsume M., Yasui K., Marumo S. J Antibiot 1989; 42: 440-448.
8. Natsume M. Ibid; 1992; 45: 1026-1028.
9. Данова С.Т., Мончева П.А., Антонова С.А., Иванова И.В. Антибиотики и химиотер 1997; 42: 4: 12-15.
10. Иванова И.В., Христова К.Р., Данова С.Т., Мончева П.А. Там же 1994; 39: 11: 21-28.
11. Tishkov S., Todorov Tz., Dundarov S., Kovachev I. Proc of FEC. 5thIntern Conf Chemistry of Nat Prod. Varna 1989; 331-335.
12. А.с. 492851/91 Тишков С., Богацевска Н. и др. 13. Kunitz M. J Gen Physiol 1946; 30: 291.
14. Мончева П.А., Данова С.Т., Антонова С.К., Иванова И.В. Антибиотики и химиотер 1997; 42: 9: 14-19.
15. Sarath G., De La Motte R.S., Wagner F.W. Proteolytic Enzymes: Practical Approach/Ed by R.J. Beynone, J.S. Bond. Oxford 1989; 25-55.
16 Kang S., Kim I., Rho Y., Lee K. Microbiology 1995; 141: 3095-3103.
17. Lowry O.H. et al. J Biol Chem 1965; 162: 420.
18. Мончева П., Ганчева А., Христова К., Иванова И. Антибиотики и химиотер. 1992; 37: 12: 21-23.
19. Laluce C., Molinari R. Biotechnol Bioeng 1979; 21: 915-938.
20. Moravkova J., Vachova L. Folia Мicrobiol 1993; 38: 201-204.
21. O'Hara M., Hageman J. J Bacteriol 1990; 172: 4161-4170.
22. Reeve C., Matin A. Ibid 1984; 160: 1041-1046.
23. Gibb G., Strohl W. Can J Microbiol 1988; 34: 187-190.
24. Ginther C. Antimicrob Agents Chemother 1979; 15: 522-526.
Написать комментарий
| 